Talán a legritkább és legkülönlegesebb színnek tartjuk a csíkos mintázatú, azaz brindle lovakat. Más fajokkal, pl. a kutyákkal ellentétben azonban lovaknál nincsen olyan gén, ami konkrétan ezért a mintázatért felelős; a brindle tehát egy fenotípus, melynek több oka lehet. Az okok közé tartozik a kevert színek egyedi megjelenése (skewed grey, skewed roan, skewed varnish roan), a fakó (dun) extrémen megjelenő primitív jegyei (extreme dun factor), a szomatikus mutáció, az incontinentia pigmenti, legeslegritkább esetben a kiméra, valamint egy újonnan felfedezett genetikai faktor, a BR1, illetve ezen eseteken túl a pigmentsejtek különböző mértékű eloszlása a Blaschko-vonalak mentén.
Láthatjuk tehát, hogy a csíkos mintázatnak számos oka lehet, épp emiatt a brindle kifejezés hasonló a fehér (white) színhez: ahogy hivatalosan nem léteznek fehér lovak, úgy hivatalosan nincs konkrét brindle mintázat sem, viszont mindkettőt (nem hivatalosan) használhatjuk gyűjtőfogalomként, egy tovább konkretizálandó fenotípus leírásaként.
Ezen felül felmerül a kérdés, hogy tudunk-e, illetve ha tudunk, megéri-e ilyen mintázatú lovakat tenyészteni?
Skewed grey, skewed roan, skewed varnish roan
Azoknál a kevert színeknél, melyeknél változó mennyiségben fehér szőrszálak keverednek a színesek közé, létrejöhet az úgynevezett skewed fenotípus, ami csíkos megjelenést okoz ezeknél a színeknél. A szürke (grey) a deres (roan), és a leopard complex gén által létrehozott varnish roan színeknél is előfordulhat ez a csíkos mintázat, a csíkok egybeesnek a Blaschko-vonalakkal (az embrionális fejlődés egy bizonyos szakaszában az embrióban intenzív sejtvándorlás megy végbe, s az epidermális sejtek vándorlásának útvonalai a Blaschko-vonalak, melyek különböző betegségek esetében szabad szemmel is láthatóvá válnak). Ha pl. a melanocyták sejtvándorlása nem megfelelően megy végbe, a Blaschko-vonalak mentén a pigmenteloszlás eltérő lesz a bőr többi részéhez képest, így jellegzetes mintázatok kialakulásához vezet. Jellegéből adódóan nem öröklődő.
A skewed roan és skewed varnish színek esetében a fehér szőrszálak ahelyett, hogy egyenletesen oszlanának el a ló testén, a Blaschko-vonalak mentén koncentrálódnak.
Reckless Dan, skewed grey quarter horse mén; későbbi élete során teljesen kiszürkült.
Catch A Bird, skewed roan angol telivér. Külön érdekesség, hogy sem a fajtában, sem felmenői között nincs jelen a roan gén, csikói közül viszont többen is örökölték tőle a deres színt: Catch A Birdnél (illetve valamelyik szülője ivarsejtjében) tehát önmagától létrejött a roan mutáció, amit képes is volt továbbörökíteni.
A fakó (dun) gén két jellegzetessége az alapszín jellegzetes kihígítása és a primitív jegyek megléte. Egyes esetekben ezek a primitív jegyek (szíjalt hát, zebroid csíkos lábak, csíkos váll) nagyon erősen expresszálódhatnak, ekkor beszélünk extrém dun faktorról, ami a brindle fenotípus egyik fajtája.
Brindle fenotípus szomatikus mutációval is keletkezhet, ekkor a Blaschko-vonalaktól eltérő mintában jelennek meg fehér csíkok az érintett egyedek testén, ahol vagy a melanocytákban keletkezett mutáció miatt nincs ezeken a részeken színtermelés, vagy a fehér mintázat kialakításában szerepet játszó területek nem az eredeti programnak megfelelően jelennek meg. (Valójában majdnem minden brindle-mintázat valamilyen mutációval keletkezett, így kicsit csalókás ilyen elnevezéssel illetni ezeket az eseteket, még ha szemmel láthatóan is különböznek a fentiektől).
DA Remote Control, arab félvér
Celano, holland melegvérű
Celano, holland melegvérű
Incontinentia pigmenti
Az incontinentia pigmenti (IP) a brindle mintázat egyetlen olyan formája, amely öröklődik, viszont mivel egy genetikai betegség, ezért sajnos egyben a leginkább olyan brindle mintázat, melyre nem érdemes tenyészteni. Nemhez (X-kromoszómához) kötött domináns mutáció, csak a kancák mutatják a brindle mintázatot és örökítik tovább, a mének még méhen belül elpusztulnak. Ahogy embereknél, úgy lovaknál is az IP egy olyan genetikai betegség, mely abnormalitásokat okoz közös fejlődési eredetük miatt nem csak a bőrön, de a fogakon, körmökön / patákon, illetve a szemeken is. Az IP olyan léziókat (elváltozásokat, sebeket) hoz létre a bőrön, melynek következtében a lovak bőrén a hyperpigmentáció mellett hypopigmentáció és szőrtelen területek is előfordulnak, s valójában ezek együttesen hozzák létre a mintát. Belátható, hogy nem egy járható út a jövőbeli brindle-tenyésztők számára.
Brenda Batty Atty, az első kanca akinél feljegyezték az öröklődő brindle mintázatot – mind ő, mind csíkos csikói incontinentia pigmentiben szenvedtek
A képen szereplő kancán jól látszik, hogy a csíkos mintázatot a betegség által okozott bőrsérülések, és a sérülések helyén fellépő szőrtelen, pigmentáltabb részek hozzák létre.
Kiméra (chimerism)
A kiméra egy nagyon ritkán előforduló jelenség, melynek során méhen belül az embrionális fejlődés korai fázisában kétpetéjű ikrek / két megtermékenyített petesejt fúzionál (összeolvad), így genetikailag mozaikos élőlény jön létre. A kiméra eredményeképpen többféle különleges mintázat is létrejöhet, melyek közül csak az egyik a brindle (a többi kiméra mintázatról majd később külön cikk fog szólni). Tehát egy nagyon ritka jelenségről van szó, s a brindle mintázatoknak ez egy ugyancsak nem tenyészthető, nem örökíthető formája.
Dunbars Gold, genetikailag tesztelt kiméra. A kiméra nemcsak a különleges brindle mintázatot eredményezte: a két embrió, ami eggyé fúzionált, egy mén és egy kanca egyed volt, így Dunbars Gold sejtjeinek egy része XX, míg másik része XY kromoszómákat hordozott (egyszerűbben mondva, egyszerre volt kanca és mén – mivel azonban a szaporító szervrendszere teljes egészében a mén-sejtvonalról származik, így ivarilag teljes értékű mén).
Tigresa Dash brindle mintázata bár elsőre extrém dun faktornak tűnik, a dun génre jellemző valódi hátszíj és egyéb primitív jegyek részleges vagy teljes hiánya (zebroid lábak, dunra jellemző csíkos váll) miatt, ami ennyire erős dun expresszió esetén elképzelhetetlen (+ elvileg felmenői között sincsen dun), megerősíti, hogy minden bizonnyal itt is kiméráról van szó.
Eltérő pigmenteloszlás a Blaschko-vonalak mentén
Ha a fentebbi eseteken túl találkozunk olyan lóval, aki egyik fentebb említett színt vagy betegséget sem hordozza, valamint nem is kiméra, és az alapszínnél sötétebb csíkokat hordoz a Blaschko-vonalak mentén, akkor a brindle fenotípus oka a fentiektől eltérő. Valamilyen oknál fogva az embriogenezis során a pigmentsejtek ezen a vonalon koncentráltabban maradtak meg, mint a test többi részén.
Brindle 1 (BR1)
2016-ban svájci kutatók publikáltak egy olyan cikket, melyben a Quarter horse fajtánál meghatároztak egy X-kromoszómához kötött gént, ami jellegzetes mintázatot okoz a lovak szőrében. A cikk alanyául szolgáló ló gazdája ragaszkodva a brindle elnevezéshez, a gén a Brindle 1 (BR1) elnevezést kapta, viszont ez a gén okozta fenotípus különbözik a többi “klasszikus” brindle mintázattól. A vertikális irányú csíkokat a csíkok helyén lévő szőrzet eltérő textúrája, esetleg eltérő pigmentsága okozza a többi normál szőrszálhoz képest. X-kromoszómához kötött szemi-domináns öröklésmenetű gén: a heterozigóta kancák, akiknél csak az egyik X kromoszómán található a BR1-mutáció, rendelkeznek a csíkos jellegű fedőszőrmintázattal, míg a homozigóta kancák és a mének (akiknél ugye csak 1 X-kromoszóma van jelen) nem mutatják ezt a mintázatot, sörényük és farokszőrzetük viszont gyérebb, mint a mutációt nem hordozó társaiké. Mivel a mintázat javarészt a szőrzet szerkezetéhez kötött, különbség adódik a hosszú téli és a rövid nyári bunda megjelenése között; télen a fedőszőr csíkjai miatt jellegzetes moth-eaten (“molyrágta”) megjelenést kölcsönöz a lónak.
A téli “moth-eaten appearance”, mellette ugyanaz a kanca nyáron, lent pedig a br1 mintázatot nem mutató mén, jól megfigyelhető gyér hosszúszőrökkel A kép a fentebb említett cikkben jelent meg.
Egyedfejlődés, mutációk: A diploid élőlényekben (azaz a lovakban, és bennünk, emberekben is) az egyedfejlődés a petesejt és a hímivarsejt egyesülésekor kezdődik. Az eleinte egysejtes zigóta folyamatos osztódásba kezd, s ebből fog később kialakulni az élőlény összes sejtje. Az egyedfejlődés korai fázisában elkülönül az ivarsejt vonal (az a sejtcsoport, amiből az ivarsejtek jönnek létre) és a szomatikus, azaz testi sejtek csoportja. A mutáció, ami az örökítőanyagban (DNS) bekövetkezett változást jelenti, mind az ivarsejt vonalban (csíravonalban), mind a szomatikus sejtekben bekövetkezhet.
A csíravonal mutációk az ivarsejt vonalban alakulnak ki, így az itt bekövetkező mutációk átadódhatnak az utódnemzedéknek is, azaz öröklődhetnek.
Amiről mi fogunk beszélni, azok a szomatikus mutációk, illetve azok egy típusa. A szomatikus, azaz testi sejtektől az ivarsejt vonal sejtcsoportja nagyon korán, az egyedfejlődés korai szakaszában elkülönülnek, így a szomatikus sejtekben bekövetkező mutációk nem öröklődő mutációk. A mutáció egyetlen sejtben történik meg az egyedfejlődés valamelyik szakaszán, azonban a folyamatos sejtosztódások során a mutáns sejt és klónjai egy (akár) látható méretű szektort / mozaikfoltot hoznak létre. Ennek a mérete különböző lehet, attól függően, hogy a mutáns sejt hány osztódáson esik át, azaz mennyire korán jött létre a mutáció az egyedfejlődés során. Az embrionális élet korai szakaszában bekövetkezett mutációk így értelemszerűen nagyobb méretű mozaikfoltot eredményeznek, mint a későbbi fázisban bekövetkezők.
Nem minden mutáció hoz létre látható változást az állatban. Ennek egyrészt az az oka, hogy a mutáció nem feltétlenül érint működő génszakaszt, vagy olyan génszakaszban következik be, amely mutációja nem okoz a biokémiai folyamatokban változást; illetve ha okoz is, diploid mivoltunk miatt a génpár másik tagja esetleges domináns jellege miatt elnyomhatja azt. Ráadásul az érintett génszakasz egyáltalán nem biztos, hogy a fenotípus kialakításáért felel. Így tehát elmondhatjuk, hogy a lovak szemmel látható szomatikus mutációi, amik például a szín megváltozását okozzák, igen ritkának számítanak.
Ha egy működő génben (amely pl. egy szín kialakulásért felel) megfelelő helyen történik a mutáció, a gén leggyakrabban funkcióvesztéses mutációt szenved, “kikapcsolódhat” a génszerkezetben történt változás miatt, és ez egy egyedi megjelenést kölcsönöz az állatnak. A cikk további részében a szomatikus mutációknak ennek a fajtáját fogom taglalni.
Az adott színekre nézve heterozigóta lovak “előnyben vannak” a szomatikus mutációkban, ugyanis ott csak egy domináns génnek kell elromolnia a mutáció következtében. Szürkék szomatikus mutációja: bloodmark, bloody shoulder A szürkékről szóló bejegyzésben már említett különleges jegyek is szomatikus mutáció következménye. A szürkeségért felelős gén (Gr) domináns gén, mely az alapszín fokozatos elvesztését, kiszürkülését okozza a rövid- és hosszúszőrökben. Ha a mutáció elrontja a Gr gént, a hibás Gr-gént tartalmazó sejt klónjai nem fognak kiszürkülni, s ez okozza a bloody shoulder és bloodmark jegyeket, ami a ló bármely testrészén megjelenhet: ezeken a területeken a ló alapszíne marad meg élete végéig, mely ugye leggyakrabban sárga vagy pej, innen adódik az elnevezés is.
bloodmark a fejen és a fülön, foltokban a test többi részén is
Stallion Cazurro II. – bár a fejen a legfeltűnőbb a szomatikus mutáció, valójában a test többi részén is jelen van
bloody shoulder almázott szürkén
Pej lovak szomatikus mutációja: A pej lovak genotípusa E_A_ (azaz EE/AA, Ee/AA, Ee/Aa lehetőségek közül valamelyik). Ha a pej színt okozó Agouti génre nézve heterozigóták (Aa), akkor a domináns Agouti allél kikapcsolódása esetén az adott genotípus E_aa lesz, ami fekete színt eredményez. A domináns Agouti-t érintő mutáció tehát egy fekete szektort eredményez a pej színű egyeden. (A mutáció ugyanígy történhetne például a domináns Extension génben is. Ha nincs működő domináns E-gén, a homozigóta recesszív ee genotípus sárga színt eredményez. Bár ez ugye sokkal nehezebben lenne észrevehető :).
Holme Park Van Gogh
Holme Park Van Gogh
Appaloosa-mintás lovak szomatikus mutációja:
Az appaloosák jellegzetes színéért és mintázataiért a leopard complex (Lp) és a pattern (PATN) gének felelősek. A Lp gén a pettyes-foltos bőr, a látható ínhártya és a sávozott paták mellett az appaloosák varnish roan-jellegét is okozza, PATN génekkel együtt pedig különböző mintázatokért felel (leopard, spotted blanket, snowcap, fewspot, stb.), allélszámtól függően.
Ha a szomatikus mutáció az Lp-gén kikapcsolását okozza, az érintett testfelületen csak a ló alapszíne dominál.
Mutáció a “színgéneken” túl:
Lorando B, Swedish warmblood mén
Lorando B, Swedish warmblood mén
Lorando B mén esete olyan értelemben egyedi, hogy a mutáns szektorról a szürke szín jut az eszünkbe – holott felmenői között nincsenek szürke lovak, így tőlük biztos nem örökölhette. Hogy lehetséges mindez?
A magyarázat az, hogy a mozaikfolt valójában csak imitálja a szürke gén tulajdonságát. A mutáció egy olyan gént érintett, melynek terméke valamilyen módon felelős lett volna a szín megjelenéséért – mivel ez a gén elromlott, a mutáns szektorban nem tud rendesen kifejeződni a ló pej színe, a rövidszőrök rendkívül kis mennyiségben tartalmaznak pigmentet.
Ez az eset egy nagyon jó példa arra nézve, hogy nem feltétlenül a színért felelős gének (Extension, Agouti) romolhatnak el. Szinte semmi esély arra nézve, hogy ezek a domináns gének egyszerre szenvedtek mutációt ugyanabban a sejtben.
Az élőlényekben a fenotípusért felelős gének megszámlálhatatlanul sok génnel működnek együtt. Ebből elég, ha csak egy elromlik, és már nem tud az adott szín kifejeződni. A konkrét színért felelős gének mellett mutációt szenvedhet bármelyik enzim génje, ami a színek előanyagait alakítják tovább, vagy elromolhatnak azok a gének, amik pl. a színlerakódásért, vagy a színtermelés valamelyik fázisáért felelnek. Minden egyes folyamathoz sok gén megfelelő működésére van szükség, elég, ha csak egy kiesik a sorból. És az alábbi lovak esetében is a fentebbiek valamelyike történhetett egy sejtben még az embrionális fejlődés egy korai szakaszában, ahonnan aztán a mutáns sejt és klónjai szemmel látható foltot, vagy épp foltokat hoztak létre.
Lorando B, Swedish warmblood mén
DA Remore Control / Clicker fehér jegyeit valószínűleg szomatikus mutáció okozhatta, de nem zárható ki a kiméra sem
A szürke szín a magyar nevezéktan szerint egy kevert szín, biológiai értelemben azonban nem szín, hanem egy pigmenttermelési defektus. Az egész állatvilágban egyedülálló, ugyanis a szürke lovak fokozatosan veszítik el alapszínüket (sokszor az úgynevezett hiperpigmentáció után kezdenek csak el szürkülni). Mindegyik szürke ló eredetileg színesen születik, és a későbbi évek során fog kiszürkülni, a szürkülés sebessége teljesen egyedfüggő, illetve valamennyire fajtafüggő is. A pigmentsejtek kimerülése a szőr színét érinti, a bőr és a szem pigmentált marad, habár ritkán a bőrön is előfordulhat pigmentvesztés, ami vitiligo formájában jelenik meg.
A szürke színt okozó gén domináns. Korábban a domináns allélt Grey (G) génként jelöltük, a legújabb kutatások azonban felfedezték, hogy több szürke-allél is létezik, attól függően, hány kópiában hordozzák a mutációt. A G2 és G3 allél nemcsak a szürkülés sebességére, hanem a melanóma kockázatára is hatással van.
A szürke árnyalatai
A szürke szín egyes árnyalatai valójában a szürkülés különböző fázisait jelentik. Az egyes árnyalatok határai természetesen szubjektívek, sokszor egybe is mosódnak, egyes fázisok akár ki is maradhatnak.
Acélszürke árnyalat akármelyik alapszín esetében előfordulhat: fekete vagy sötétpej lovak kiszürkülésénél szinte mindig megfigyelhető, ám a világosabb színű (pej, sárga, fakó, stb.) lovak a hiperpigmentáció miatt teljesen besötétedhetnek a kiszürkülés előtt, így az acélszürke árnyalat náluk is ugyanúgy jelen lehet. Részben emiatt a szürkülés egy bizonyos fázisa után már nem lehet megállapítani az eredeti alapszínt, hiszen az acélszürke nem jelent automatikusan egyet a fekete vagy sötétpej alapszínnel.
Acélszürke
Acélszürke
Mézszürke árnyalat akkor jelentkezik, amikor a sárga, pej ló (még) nem mutat drasztikus hiperpigmentációt, viszont a fokozatos pigmentvesztés egyfajta mosott színt eredményez. (Elvileg nem lehetetlen árnyalat fekete alapszín esetében sem, hiszen a fekete pigmentek hígulása is bronzos árnyalatot eredményez).
Mézszürke
Mézszürke
A szürkülés következő fázisa az almázott szürke, amikor a különböző sejtek pigmentvesztése nem egyenletes a bőr alatt futó kapillárisok (vérerecskék) miatt: az almázott mintázat ezeknek a vérerecskéknek a mintázatát követi. Helyesen ők az almázott szürkék, és nem almásderesek. Almázott szürke állapotban már az eredetileg világosabb színű lovak (sárgák, pejek) is sötétbarna-szürke-fekete árnyalatú szőrökkel rendelkeznek a hierpigmentáció miatt, így egy almázott szürke esetében már biztosan nem lehet még betippelni sem az eredeti alapszínt.
Mézszürke és almázott szürke közötti átmenet
Almázott szürke
A teljesen kiszürkült ló neve ezüstszürke, a laikusok gyakran ezeket a lovakat hívják tévesen fehérnek.
Ezüstszürke
Ezüstszürke
A legyesszürke / szeplősszürke / pisztrángszürke a szürkülés végső, és egyetlen olyan fázisa, ami után a ló a kifehéredés helyett egyre színesebb lesz. A kimerült pigmenttermelő sejtek újra elkezdenek melanint (pigmentet) termelni, így a ló pöttyökben visszaszínesedik, az idő előrehaladtával egyre jobban.
A domináns szürke mutáció túltermelteti a pigmenttermelő sejteket, amik előbb-utóbb kimerülnek, és nem lesznek képesek pigmenttermelésre. A pigment túltermelés következménye a hiperpigmentáció, ami csikók esetében a legfeltűnőbb, de a kiszürkülés végleges megkezdése előtt is jelen lehet. A szürke csikók bár színesen születnek, a hiperpigmentáció miatt jóval sötétebbek, mint a nem szürke csikók, így ennek, illetve a szem környékén megfigyelhető szürke elszíneződésnek, “szemüvegnek” (grey goggles) köszönhetően a később kiszürkülő csikók az esetek egy részében jól megkülönböztethetők a nem kiszürkülő csikóktól.
Hiperpigmentált csikó szürke “szemüveggel” (sárga vagy pej alapszín)
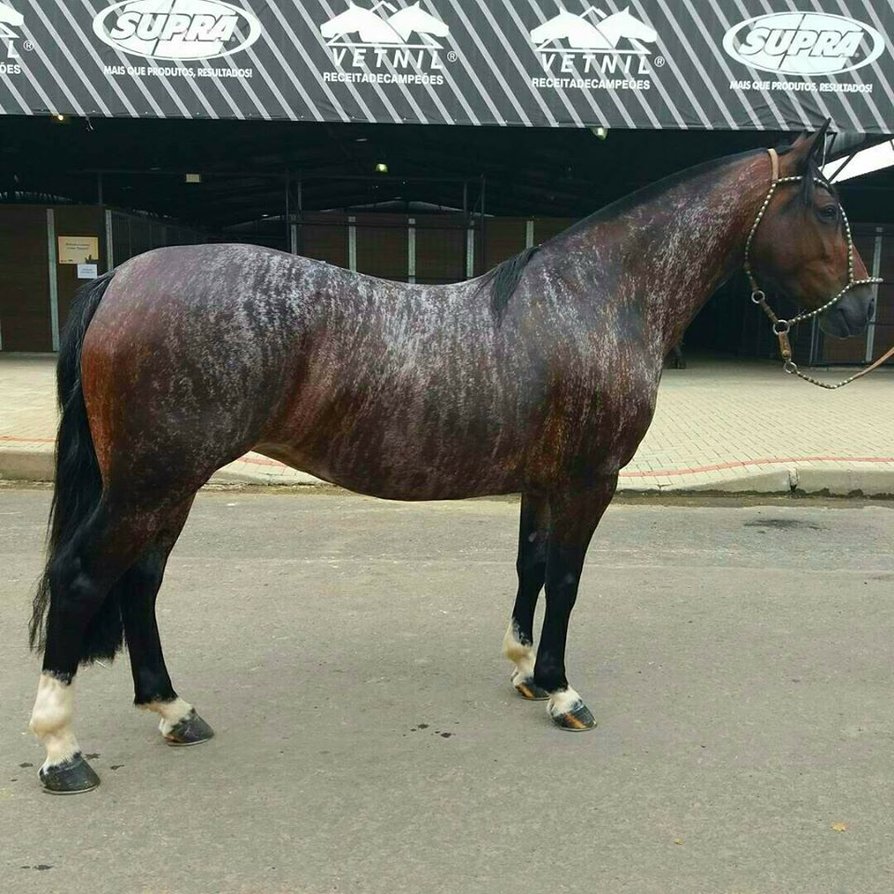
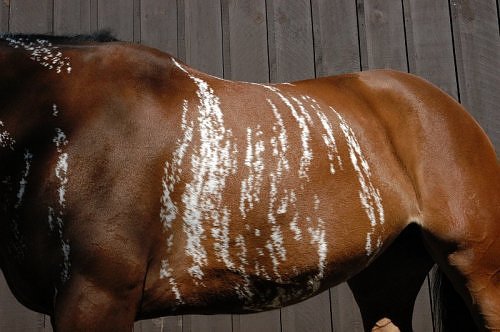
A kiszürkülés előtt sok világos színű ló színe (sárgák, pejek, palominók, stb.) előbb besötétedik, és ebből a sötét színű árnyalatból fognak majd kivilágosodni. Ezért a szürkülés egy bizonyos fázisától (acélszürke, almázott szürke) már képtelenség megállapítani a ló eredeti alapszínét.
Szürkülő palomino – minden “lovas színgenetikus” kedvenc fotója a
kiszürkülés előtti hiperpigmentáció mértékéről, avagy miért nem
tippelünk alapszínt almázott szürkék esetében
A szürkülés (szürke szálak megjelenése) leghamarabb a fejen, illetve a farok végén kezdődik, aminek a vége így mosottas jelleget kap. A kiszürkülés módja, sebessége lófüggő. Vannak lovak, akiknek a teste sokkal hamarabb szürkül, a hosszúszőrök pedig nagyon sokáig sötét színűek maradnak, míg másoknál éppen ellentétesen a szürkülés, a sörény és farok szinte már fehér, míg a ló teste még acélszürke árnyalatú. Mindkét esetben a sötét testrészek is ki fognak szürkülni, előbb vagy utóbb. A szürkülés részben szürke szőrszálak megjelenéséből (ez általában a fejen a leglátványosabb), részben pedig a testen lévő pigmentek fokozatos kihígulásából áll, ezért sok szürkülő ló színe lesz olyan furcsa, barnás, színvesztett, a mézszürke árnyalat is így jön létre.
A szürkülés minden lóban máshogy zajlik le, és néha nagyon különleges színek jönnek létre. Tetrarch spots vagy chubari spots jellegnek nevezzük azt, amikor a ló testén kör alakú foltokban gyorsabb a kiszürkülés, mint a test többi részén. A foltok átmenetiek, a többi rész fokozatosan hozzászürkül, így a foltok előbb-utóbb eltűnnek.
Chubari vagy Tetrarch-spots
Szomatikus mutációs folt akkor következik be, amikor egy foltban a mutációt okozó génszakasz elvész vagy kikapcsol, a mutáció hatása megszűnik, így az érintett folt nem fog kiszürkülni. Maradandó jelleg, ahogy a ló egyre világosabb, úgy lesz egyre kontrasztosabb a mutációs folt. A legtöbb esetben éles határvonalú, erősen pigmentált (=a ló eredeti színével megegyező) folt.
Szomatikus mutáció a fejen
Szomatikus mutáció a fejen
Bloodmark vagy bloody shoulder esetében a pigmenttermelő sejtek egy foltban egyszerre kapcsolnak vissza pigmenttermelésre. A mechanizmus ugyanaz, mint a legyesszürke szín esetében, csak apró, különálló foltok helyett egybefüggő terület lesz újra pigmentált. Általában hamar bekövetkezik, így sokszor nehéz megkülönböztetni a szomatikus mutációtól. A szomatikus mutációval ellentétben nem éles határvonalú, hanem szaggatottabb, foltos jellegű, kevésbé erős színű, mint a szomatikus mutációs folt.
Bloodmark/bloody shoulder
Valószínűleg Bloodmark/bloody shoulder, de nem elképzelhetetlen, hogy valójában szomatikus mutáció
Gyakran összekeverhető más színekkel
A szürkéket gyakran keverik a deres, a fehér, és az appaloosák varnish roan színével.
A fentebbi linkekre kattintva elérhető az adott színekről és génekről szóló részletes és illusztrált leírás.
Nem szürke, nem is albínó vagy cremello: “fehér”, pontosabban White spotting 14 mintázatú angol telivér
Nem szürke; vasderes
Nem szürke; varnish roan kismértékű spotted blanket mintával
Heterozigóták vs. homozigóták
Bár a szürke mutáció domináns öröklésmenetű génnek tekinthető, allél-dózis hatás van jelen, azaz nem mindegy, milyen és mennyi szürke-allélt hordoz egy ló. Így nemcsak a hetero- és a homozigóták között figyelhető meg a különbség, hanem az egyes G2-G3 allélok között is.
A heterozigóta, lassan szürkülő G2 szürkék (G2/G1, vagy G2/N, akik csak az egyik szülőtől örökölték a génváltozatot, és csak 50% eséllyel örökítik), gyakrabban lesznek legyesszürkék, a szomatikus mutáció és bloodmark jegyek is gyakrabban figyelhetők meg náluk. Ennek oka, hogy a G2 mutációt, ami egy duplikáció, csak az egyik szülőtől örökölték, így ez a plusz génszakasz csak egy példányban van meg, nagyobb eséllyel veszhet el/inaktiválódhat, mint homozigóták esetén vagy mint a három kópiát is hordozó G3 lovak esetében.
G3/G1 heterozigótáknál, valamint a többi génvariáció esetén is (G2/G3, G2/G2) a szürkülés sebessége gyorsabb, hamarabb érik el az ezüstszürke színárnyalatot, ritkábban lesznek legyesszürkék, és akkor is kevesebb “szeplővel” rendelkeznek. Hajlamosabbak a vitiligo-ra (a bőr foltokban történő pigmentveszteségére), valamint a mutáció okozta melanóma is nagyobb eséllyel jelenhet meg náluk (bár a melanóma gyakorisága, lefolyása további tényezőktől is függ). A G3/G3 homozigóták extrém gyorsan szürkülnek, melanóma kockázatuk igen magas.
Vitiligo
Vitiligo
Melanóma
A szürke színt okozó mutáció jellege miatt egyes gének túlműködnek, ami így a pigmenttermelő sejtek daganatához, melanómához vezet. Az ezzel kapcsolatos publikációk szerint a tizenöt év feletti szürke lovak 70-80%-ánál kialakul, és az esetek nagyobb részében a kezdetben jóindulatú daganat rosszindulatúvá alakul, és áttéteket, metasztázisokat képez. Azóta kiderült, hogy úgy tűnik, erősen fajtafüggő a dolog, az újabb kutatások szerint az Andalúzoknál a leggyakoribb, ezt követik a Lipicai lovak, míg például a Quarter Horse fajtában kifejezetten kevés szürkét diagnosztizáltak melanómával. A legújabb kutatások pedig további magyarázattal szolgálnak a jelenségre, a melanóma kockázata ugyanis erősen függ a mutáció kópiaszámától, azaz hogy egy szürke G2 vagy G3 alléllal rendelkezik-e, és milyen zigozitású. Ráadásul, genetikai okok miatt gyakrabban alakulhat ki olyan homozigóta szürke lovaknál, akik fekete alapszínnel rendelkeznek.
A melanóma leggyakrabban a szőrtelen részeken jelenik meg (farok alatt, illetve a fejen), és az emberi melanómával ellentétben nem befolyásolja a kialakulás mértékét vagy esélyét az UV, ugyanis nem külső hatás indukálta daganat, hanem genetikai eredetű.
További képek
steel grey – acélszürke (internetes kép)
rose grey – mézszürke (pej alapszín) (internetes kép)
rose grey – mézszürke (sárga alapszín) (internetes kép)
Mazsola – szürke arab félvér spotted blanket mintával (Pintér Csilla)
Sheba, 75% andalúz 2011-ben és 2016-ban (Dunai Cynthia)
Sally – araappaloosa (Kassai Fanni)
light grey – ezüstszürke (internetes kép)
Lucifer – shagya arab 2014-ben és 2016-ban (Perlaky Alexa)
Deepforest Grizzly – legyesszürke shagya arab x quarter horse (Szilágyi Zsófia)
Gréti – legyesszürke arab félvér (Pintér Csilla)
Gandalf – almázott- & legyesszürke osztrák melegvérű (Lendület Lovasiskola)








































 Szürkülő palomino – minden “lovas színgenetikus” kedvenc fotója a
Szürkülő palomino – minden “lovas színgenetikus” kedvenc fotója a

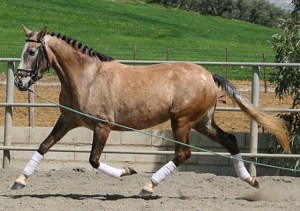
 Chubari vagy Tetrarch-spots
Chubari vagy Tetrarch-spots


























