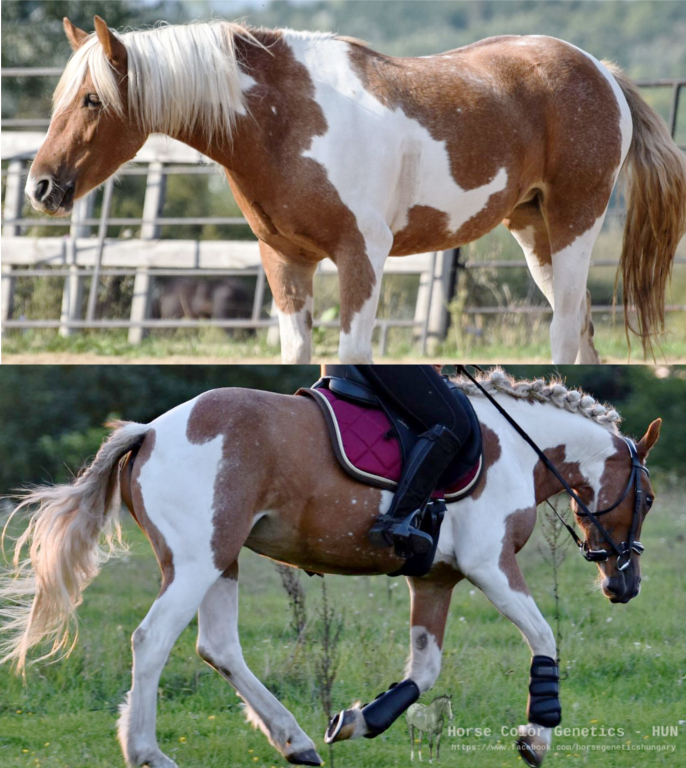
Leopard complex és varnish roan
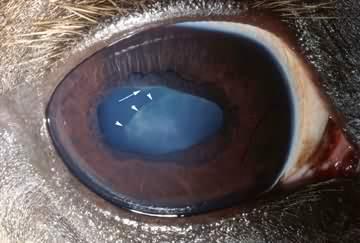
- a fehér ínhártya (“emberi szem”)
- a sávozott pata
- a pettyes (foltokban depigmentálódott) bőr, ami a legfeltűnőbb a száj, szem, genitáliák környékén
- a kor előrehaladtával egyre nagyobb mértékben jelentkező (=progresszív) depigmentáció, “lisztes” mintázat, a varnish roan




A varnish roan egy depigmentáció, ami a kor előrehaladtával egyre nagyobb mértéket ölt (progresszív; ellentétben az ismert fehér mintázatokkal), szimmetrikus, és a csípő fölötti területről indul. A mutáció által létrejött depigmentáció azzal jár, hogy a bőrben a melanociták (pigmenttermelő sejtek) száma csökken, így mind a bőrben, mind a szőrben pigmenthiány lesz jelen. Ránézésre tipikusan olyan, mintha a lovat liszttel borították volna be. A pigmentveszteség akár extrém módon is jelen lehet, szinte teljesen “kiszürkítheti” a lovat, anélkül, hogy jelen lenne a szürkeséget okozó gén. A varnish roan további jellegzetessége, hogy a csontos felületek (orrhát, lábvégek, ízületek) pigmentáltak maradnak (hőérzékeny allél-jelleg), így egy nagyon egyedi mintázat jön létre.
A Leopard complex egy “kapcsológén”: minden más appaloosa-mintázat (foltos, pettyes, takaró, stb.) további módosító génekkel jön létre, ám csak abban az esetben, ha az Lp is jelen van. Ha nincs meg az Lp, akkor hiába hordozza egy ló ezeket a mintázatgéneket, azok semmiféle különleges jelleget nem fognak okozni.
Vannak különleges varnish roan mintázatok, amikor a varnish roan nem “lisztszerűen” borítja be a lovat; ilyen mintázat pl. a snowflake, amikor a depigmentáció fehér pöttyökben jelenik meg (hópehely-jelleg). Egyes spotted leopard (lsd. lentebb) lovaknál a pöttyök körül halozóna jelenhet meg, ezt a mintázatot peacock-appaloosa-nak nevezik (vagy: leopard appaloosa with peacock spots).




Pattern gének, PATN1
A Leopard complex létrehozza tehát az “appaloosa”-jegyeket és a varnish roan mintázatot önmagában, valamint lehetővé teszi, hogy további mintázatok alakulhassanak ki. Minden egyéb appaloosa-mintázatot (leopard, spotted blanket, stb.) további gének, ún. mintázatgének (pattern gének) hoznak létre. Többféle pattern gén van, jelenleg egyet sikerült genetikailag azonosítani, illetve genetikai színtesztelni, ez a PATN1 (pattern1) gén.
A Leopard complex inkomplett (azaz nem teljes) domináns jellegének leginkább itt van jelentősége. A PATN1 szimpla domináns gén, mindegy, hogy egy vagy kettő domináns PATN1 alléllal rendelkezik-e a ló, nincs különbség homo- és heterozigóta PATN1 génes lovak között. Az azonban nem mindegy, hogy az Lp génre nézve homo- vagy heterozigóta-e a ló.
Heterozigóta Lp mellett alakulnak ki a tipikusan pöttyös minták, amikor a lovon kisebb vagy nagyobb méretű, de mindenképpen több darab pötty van jelen. Ilyen mintázat pl. a spotted leopard vagy leopard appaloosa, illetve a spotted blanket.











CSNB, Congenital Stationary Night Blindness
A fenotípus és a genotípus kapcsolata
Az Lp inkomplett domináns jellege miatt fenotípusosan is megkülönböztethetők a heterozigóta és homozigóta Lp-lovak.
Ha csak a Leopard complex van jelen, a fenotípusos megkülönböztetés nem 100%-os, ám ebben az esetben is vannak különbségek:
– a homozigóta Lp/Lp lovak patája inkább viaszsárga, az appaloosa jellegek erősebben vannak jelen (jobban depigmentálódott bőr, erősebb varnish roan jelleg), CSNB jelen van
Leopard complex + PATN1 esetében:


- Lp/Lp (homozigóta) lovaknál megfigyelhető appaloosa-mintázatok PATN gének mellett
Így tehát szemre is megállapítható, hogy a feltűnő, sok pöttyel rendelkező lovak egészen biztosan heterozigóták (nem 100%-os színörökítők); míg a fewspot, vagy pötty nélküli takaróval (snowcap) rendelkező lovak homozigóták, legalább a Lp-gént 100%-ban örökítik.
Progresszív varnish roan egyéb meglévő appaloosa-minták mellett is jelen lehetnek: míg egyes lovak mintázata (pontosabban a depigmentálódott területe) “fix”, addig más lovaknál a varnish roan fokozatosan növeli a mintázat körül a fehér (depigmentált) területet.

Lp color shifting


Felhasznált irodalom:
Rebecca R. Bellone et al.: Differential Gene Expression of TRPM1, the Potential Cause of Congenital Stationary Night Blindness and Coat Spotting Patterns (LP) in the Appaloosa Horse (Equus caballus); Genetics 179: 1861–1870 (August 2008)
H. M. Holl et al.: Variant in the RFWD3 gene associated with PATN1, a modifier of leopard complex spotting; Animal Genetics, 47, 91–101 (August 2015)
Rebecca R. Bellone et al.: Fine-mapping andmutation analysis of TRPM1: a candidate gene for leopard complex (LP) spotting and congenital stationary night blindness in horses; Oxford University Press (2010)
Rebecca R. Bellone et al.: Evidence for a Retroviral Insertion in TRPM1 as the Cause of Congenital Stationary Night Blindness and Leopard Complex Spotting in the Horse; PLOS ONE, 8 (October 2013)
R. B. Terry et al.: Assignment of the appaloosa coat colour gene (LP) to equine chromosome 1; Animal Genetics, 35, 134–137 (2004)